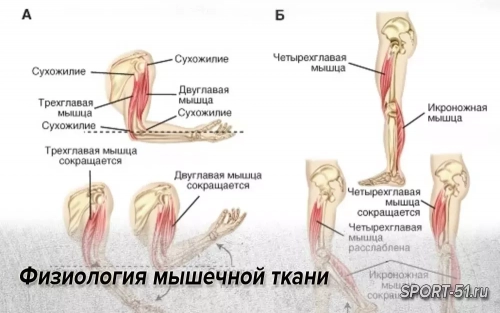
Сердечная мышца обладает некоторыми характеристиками скелетной мышцы и некоторыми характеристиками гладкой мышцы. Из-за вязкости крови сокращения сердечной мышцы должны быть достаточно продолжи тельными, чтобы опустошить камеры сердца.
Сердечная мышца обладает некоторыми характеристиками скелетной мышцы и некоторыми характеристиками гладкой мышцы. Из-за вязкости крови сокращения сердечной мышцы должны быть достаточно продолжи тельными, чтобы опустошить камеры сердца.
Энергия запасается в эластичных стенках сосудов, благодаря которым кровь постоянно двигается по сосудам. Необходимо помнить, что ритмические сокращения скелетной мускулатуры также важны для циркуляции крови. Сердечная мышца обладает уникальным аэробным обменом. Способность получать энергию через анаэробный гликолиз в этой ткани ограничена.
В состоянии покоя потребление кислорода на один грамм сердечной мышцы больше, чем потребление кислорода скелетной мышцы при тяжелой физической нагрузке. При физических нагрузках коронарный кровоток возрастает до 4-х раз, а сердечная мышца обладает уникальной способностью выделять кислород из крови.
На сердце лежит огромный объем работы. Подсчитано, что около 20% энергии в нормально работающем желудочке тратится на работу, не связанную с сокращением. Около 70% энергии тратится на механическую работу левого желудочка. Он работает автоматически и сокращается около одного раза (три раза во время тяжелой физической нагрузки) в секунду.
Таким образом, его потребление кислорода даже в состоянии покоя в 4-5 раз больше, чем в скелетной мышце. Артериовенозное различие в давлении кислорода также выше. Основная часть объема клетки (20-30%) - это митохондрии, в то время как в остальных мышцах они занимают меньше 10% объема.
Поскольку сердечная мышца работает непрерывно, ее способность потреблять различные вещества меняется. Сердечная мышца обладает уникальными метаболическими характеристиками. При обмене веществ в состоянии покоя 70% потребности сердечной мышцы в энергии поставляется из жирных кислот.
Свободные жирные кислоты, также как продукты их превращения, такие как ацетоуксусная кислота и бета-гидроксимасляная кислота, которые в большом количестве выделяются из печени при голодании, очень подходящие для использования вещества. Однако существуют различия в удобстве потребления среди жирных кислот из-за, например, активности карнитинпальмитоилтрансферазы.
Удобными веществами являются триацилглицерины, образованные в печени из липидов крови, также как цикломикроны, синтезированные в кишечной слизистой оболочке. Роль глюкозы как энергетического запаса небольшая, а большое количество циркулирующих жирных кислот замедляет метаболизм глюкозы в сердечной мышце.
Инсулин и глюкоза стимулируют гликолиз и подавляют окисление жирных кислот. Однако в диабетическом состоянии наблюдается внезапный спад в окислении лактата, по крайней мере, у крыс. Замедление цикла трикарбоновых кислот приводит к быстрому окончанию сокращения, что и следовало ожидать. Митохондриальный переносчик аденина-один из регуляторных сайтов в дыхательном контроле сердечной мышцы.
Из-за того, что сердце - это полый орган, и для перекачивания необходимо, чтобы различные клетки взаимодействовали между собой (сеть кардиомиоцитов), кардиоциты связаны друг с другом мостиками с низким сопротивлением, которые позволяют продвигаться волне сокращения. Из-за непрекращающихся циклов сокращений периоды отдыха короткие. Наличие абсолютного рефракторного периода в каждом цикле защищает сердечную мышцу от истощения. В этот период сокращение не может быть остановлено.
Перекачивание ионов кальция происходит по нескольким механизмам. Поскольку кальций необходим для сокращения, он проникает в клетки сердца как через плазматическую мембрану в цитоплазму, так и из саркоплазматического ретикулума. Было подсчитано, что более 10% энергии в нормально работающем левом желудочке потребляется во время работы Na+/K+ и Са++ АТФаз в процессах возбуждения и электромеханического сопряжения. Ионы кальция также стимулируют окислительное фосфорилирование помимо способности к сокращению. Нарушения в цитозольном транспорте кальция играют важную роль в опосредовании сократительной дисфункции при сердечном ударе.
Сердечные заболевания встречаются очень часто, а коронарная недостаточность сердца -одна из наиболее частных причин смерти в западных странах. Хотя метаболизм сердечной мышцы аэробный, в анаэробном и ишемическом состоянии, когда кислород недоступен для сердечной мышцы, мышца получает энергию из анаэробного гликолиза.
Значительно повышенный уровень гликолиза при ишемии сопровождается высоким потреблением глюкозы и повышенным образованием молочной кислоты. Предполагалось, что это является причиной болей в груди при ишемии. Однако перемежающаяся умеренная ишемия может привести к согласованному клеточному ответу, включая повышение уровня аэробного обмена глюкозы, изменению экспрессии сердечных генов и развитию специфических механизмов для программируемого выживания клеток (подготовка, «оглушение» и «спячка»).
Около 95% энергии, потраченной в сердечных мышцах, используется для синтеза АТФ в митохондриях. При тяжелой ишемии в результате распада АТФ образуется АДФ, АМФ и аденозин. Аденозин способен проникать через оболочку сердечной мышцы, а его высвобождение при ишемии ведет к полной его потере.
Ишемия, продолжающаяся 30 мин, уменьшает запас аденозина в клетках сердечной мышцы на 50%. Запасание аденозина ~ очень медленный процесс: за один час только 2% аденозина может быть заново синтезировано. Серьезное истощение запасов аденозина и АТФ является основной причиной смерти при сердечной ишемии.
Гипоксия также приводит к повышению количества белков теплового шока в миокарде, по крайней мере, у животных. Было показано, что фруктозо-1,6-дифосфат защищает сердечную ткань при гипоксии, как и при изолированной сердечной перфузии. КФ является также важным способом переноса химической энергии из митохондрий и хранилищем макроэргического челнока в сердечной мышце.
Слабеющий миокард характеризуется уменьшением запасов КФ и общего содержания креатина и понижением уровня белка-транспортера креатина.