 Кислород транспортируется по дыхательным путям к месту окисления в активных тканях. При интенсивных нагрузках активные клетки скелетной мышцы устанавливают потребность в кислороде, поскольку более 90% энергии тратится на мышцы.
Кислород транспортируется по дыхательным путям к месту окисления в активных тканях. При интенсивных нагрузках активные клетки скелетной мышцы устанавливают потребность в кислороде, поскольку более 90% энергии тратится на мышцы.
Кислород связывается с гемоглобином в эритроцитах, в то время как субстраты перемещаются в плазму. Поставка кислорода должна быть непрерывной, поскольку в организме большинства млекопитающих имеется лишь минимальный запас кислорода, а субстраты хранятся как в мышечных клетках, так и в тканях в большом количестве.
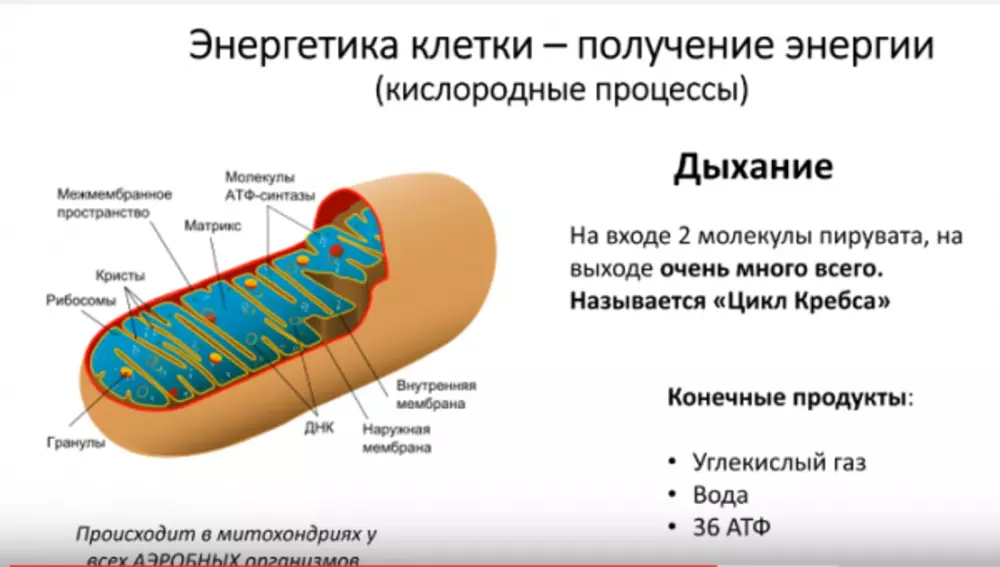
Окислительное фосфорилирование в митохондриях
Аэробный энергетический обмен происходит в митохондриях и приводит к наибольшему выходу энергии. Как следует из названия, для этого процесса требуется кислород. Аэробный гликолиз - наиболее эффективный способ образования энергии. В анаэробных условиях только 2 моль АТФ синтезируется из 1 моль гликогена, в то время как в присутствие кислорода 38 моль АТФ может синтезироваться из того же количества гликогена.
Чистый выход АТФ 36 моль, за вычетом потраченных 2 моль АТФ (38 молекул в сердечной мышце, столько же в почках и печени). В митохондриях мышечных клеток атомы водорода выделяются из восстановленных веществ, которые образуются в ходе цикла трикарбоновых кислот, в процессе, называемом окислительным фосфорилированием. Специальные белковые комплексы, переносящие электроны, передают электроны от атома водорода молекуле кислорода.
Энергия, высвобождаемая во время транспорта электронов, сохраняется в виде АТФ. Электроны по дыхательной цепи переносятся в порядке повышения окислительно-восстановительного потенциала -от более электроотрицательного вещества к более электроположительному кислороду.
 В клетках при высоком парциальном давлении кислорода потребность в энергии определяет уровень митохондриального дыхания, а снабжение субстратами определяет энергетический уровень клетки, при котором достигается этот уровень митохондриального дыхания. Запас АТФ и КФ мало сравним с количеством энергии, которая требуется для активной мышечной работы.
В клетках при высоком парциальном давлении кислорода потребность в энергии определяет уровень митохондриального дыхания, а снабжение субстратами определяет энергетический уровень клетки, при котором достигается этот уровень митохондриального дыхания. Запас АТФ и КФ мало сравним с количеством энергии, которая требуется для активной мышечной работы.
Из-за малого запаса макроэргических фосфатов АТФ может тратиться больше или меньше, чем синтезироваться, лишь в небольшие промежутки времени. Таким образом, синтез АТФ должен происходить, когда обычно клеточные процессы гидролизуют АТФ. Поэтому митохондриальное окислительное фосфорилирование тесно связано с несколькими обменными путями и быстро отвечает на изменения в клеточных потребностях в АТФ. Однако связь дыхательной цепи с другими обменными процессами не столь высокая.
Было подсчитано, что около 1~3% кислорода, поступившего в дыхательную цепь в состоянии покоя, выходит из нее в виде пероксида. Образование высокореакционноспособных частиц кислорода может иметь некоторое влияние, как в состоянии покоя, так и при физических нагрузках.
Митохондрии в скелетных мышцах потребляют наибольшее количество кислорода и служат основным источником энергии в обменном процессе для поддержания нормальной работы. Цикл лимонной кислоты (также называемый циклом Кребса или циклом трикарбоновых кислот) - это ряд реакций в митохондриях, которые приводят к распаду ацетил-КоА, высвобождая восстановительные эквиваленты, которые используются в окислительном фосфорилировании для образования АТФ из АДФ.
Цикл трикарбоновых кислот играет основную роль в аэробных обменных процессах. Он является заключительным путем метаболизма глюкозы (гликолиз), жиров (бета-окисление жирных кислот) и белков. Как в состоянии покоя, так и при физических нагрузках бета-окисление жирных кислот и гликолиз предоставляют более 95% ацетил-КоА, вступающего в цикл трикарбоновых кислот.
В цикле лимонной кислоты ацетил-КоА соединяется с оксалоацетатом, формируя соль лимонной кислоты. В последующих реакциях оксалоацетат снова регенерируется. В результате выделяется 3 НАД Н, 1 ФАД Н2, 1 ГТФ и 2 С02. Далее восстановительными эквивалентами подвергаются окислительному фос-форилированию. 3 НАД Н и 1 ФАД Н2 дают в результате 12 АТФ.
Рис. Распад глюкозы с выходом АТФ 36-38 моль посредством гликолиза, цикла Кребса и электроннотранспортной цепи






